Table of Contents
Definition
noun
plural: nuclear envelopes
nu·cle·ar en·ve·lope, ˈn(j)ukliɚ ˈɛn.və.ləʊp
The two layered membrane that encases the nucleus of a eukaryotic cell, which separates the nucleus from other cell organelles
Details
Overview
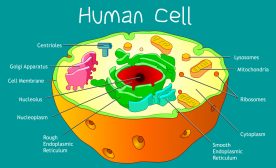
The cell nucleus is the organelle of the eukaryotes responsible for maintaining the integrity of DNA and for controlling cellular activities such as metabolism, growth, and reproduction by regulating gene expression. It is a double-membraned organelle and this double membrane is referred to as nuclear envelope (also called nuclear membrane, plasmalemma, sor karyotheca). The nuclear envelope surrounds most of the eukaryotic cell’s genetic material as opposed to the genetic material of the prokaryotes that is not membrane bound.
Features
The nuclear envelope is a lipid bilayer. It has nuclear pores that control the movement of molecules between the nucleoplasm and the cytoplasm. It is impermeable to large molecules. Thus, it separates the contents of the nucleus from the cellular cytoplasm and allows entry of selected molecules.
In between the two membranes of the nucleus is a space called the perinuclear space. The outer membrane is contiguous with the endoplasmic reticulum. The inner membrane is constituted by a network of filaments called nuclear lamina. The lamina attach to chromosomes. It also acts like a shield for the nucleus. The outer and the inner membranes are connected by the nuclear pores. In mammals, the nuclear envelope spectrin repeat proteins are expressed in the outer membrane. These proteins connect the cytoskeletons in the cytoplasm to the nucleoskeleton in the nucleoplasm. This connection aids in the positioning of the nucleus within the cell.
Common biological reactions
Common biological reactions
During cell division, the nuclear envelope undergoes major changes in animal and plant cells. The nuclear envelope disintegrates to allow the spindle fibers to access the chromosomes in the nucleus. An exception to this is the yeast cells whereby the nuclear envelope stays intact during cell division. In animal and plant cells, the nuclear envelope breaks down into pieces during prometaphase of mitosis. Then, during telophase, the nuclear membrane reforms. The reformation process remains unclear how it proceeds. It is postulated to occur via vesicle fusion or probably by reshaping of the endoplasmic reticulum, enclosing the nuclear region with a new nuclear envelope.
Common biological reactions
The presence of the nuclear envelope prevents the easy passage of large molecules (e.g. proteins and RNA) into and out of the nucleus. Although the nuclear envelope is perforated with nuclear pores, large molecules would still need a nuclear transport mechanism in order to enter and exit the nucleus. Small molecules, such as ions, can pass through the nucleus with ease. However, cargo proteins and RNAs that need to be transported require importins and exportins to enter and exit the nucleus, respectively. On one hand, the cargo binds with the importin in the cytoplasm, and then moved into the nucleus through the nuclear pore. On the other hand, the cargo binds with the exportin inside the nucleus, and then moved outside the nucleus via the nuclear pore. Nuclear transport needs energy to proceed. Thus, GTPases (e.g. Ran enzyme) help by hydrolyzing GTP (guanosine triphosphate) so that energy would be released in the process. The energy released would be used to dissociate the cargo from the importins and to bind the cargo to the exportins.
Biological functions
The nuclear envelope compartmentalizes the nucleoplasm, setting boundaries between the nucleus and the cytoplasm. Nevertheless, it is perforated with holes called nuclear pores that regulate the exchange of substances (for example, proteins and RNA) between the nucleus and the cytoplasm. The nuclear transport of the large molecules like proteins and RNAs occurs via an active transport system carrier proteins while the passage of small molecules and ions occur passively via the nuclear pores.
Supplementary
Etymology
- origin: word (translation, “meaning”)
Synonym(s)
Further reading
See also
© Biology Online. Content provided and moderated by Biology Online Editors